


Autor: MVZ. DEAPP. Guadalupe Edgar Beltrán-Rosas
Technical Director in Swine
Institución: ebios nutrition S.A.P.I. de C.V.
https://orcid.org/009-0002-8141-4979
beltran@ebios-nutrition.com
![]()
- Introducción
El circovirus porcino (PCV por sus siglas en inglés; porcine circovirus) se describió por primera vez en 1974 por Tischer y colaboradores como un agente contaminante de la línea celular PK-15 obtenida a partir de riñón de cerdo. Este virus se consideró inicialmente como no patógeno. Un virus similar a PCV fue aislado por primera vez en cerdos con enfermedad debilitante en el oeste de Canadá en 1991, en granjas de alto estatus sanitario, se identificó una variante de circovirus porcino el cual fue aislado de cerdos afectados por el síndrome del desmedro multisistémico postdestete (PMWS, por sus siglas en inglés: postweaning multisystemic wasting syndrome). Poco tiempo después, un virus prácticamente idéntico al aislado canadiense fueron detectados en cerdos enfermos en Estados Unidos y Europa. La secuenciación de los dos virus mostró una divergencia genética significativa y de ahí que posteriormente el Comité Internacional de Taxonomía de los Virus sugirió llamar al PCV no patógeno circovirus tipo 1 (PCV-1) y al nuevo virus asociado con la enfermedad clínica como circovirus porcino tipo 2 (PCV-2).
El PMWS se considera una enfermedad multifactorial, incluidos otros agentes infecciosos y el estado inmunológico del animal, siendo el circovirus porcino tipo 2 (PCV-2) el agente imprescindible para su desarrollo, una afección caracterizada por pérdida de peso, retraso en el crecimiento, una reducción de la eficiencia alimenticia, dificultad respiratoria y lesiones del tejido linfoide, lo que resulta en una menor productividad y rentabilidad. Dicho síndrome se conoce actualmente como enfermedad sistémica por PCV-2 (PCV-2-SD, por sus siglas en inglés: systemic diseases) o circovirosis porcina (CP), dicha terminología esta designada específicamente al PMWS.
- Implicaciones económicas
La circovirosis porcina (CP) es una enfermedad de distribución mundial que ha causado una pérdida económica considerable, causada por el circovirus porcino tipo 2 (PCV-2). Es el agente principal de la enfermedad asociada al circovirus porcino (PCVAD, por sus siglas en inglés: porcine circovirus-associated diseases). Las implicaciones económicas son multifacéticas y afectan varios aspectos de la industria porcina. Las infecciones por PCV-2 también hace que los cerdos sean más susceptibles a patógenos secundarios como el virus del síndrome reproductivo y respiratorio del porcino (PRRSV, por sus siglas en inglés: porcine reproductive and respiratory syndrome virus), el Parvovirus porcino (PPV, por sus siglas en inglés: Porcine Parvovirus), Mycoplasma spp, por inmunosupresión, lo que resulta en un aumento de la mortalidad. La PCVAD incurre en un costo promedio de 3 – 4 USD (hasta 20 USD) por cerdo en los Estados Unidos y, por lo tanto, es reconocido como un patógeno económicamente importante en la industria porcina mundial.
Cabe mencionar, que los costos asociados con el diagnóstico, el tratamiento y la prevención de las infecciones por PCV-2 aumentan la carga económica. Además de ello, debido a las infecciones silentes con signos inaparentes (infección subclínica) que merman las tasas de conversión, y a la producción de cerdos no comercializados causando importantes pérdidas económicas en un ámbito tan intensificado como es hoy en día la crianza moderna del porcino.
Los esfuerzos para controlar la propagación del PCV-2 requieren inversiones en medidas de bioseguridad, prácticas de higiene y programas de vacunación. El virus es muy resistente en el ambiente, mostrando también una gran resistencia a los tratamientos químicos y térmicos. El PCV-2 es relativamente resistente, bajo condiciones in vitro, a diferentes desinfectantes comerciales a base de clorhexidina, formaldehido, yodo o alcoholes. Se ha demostrado que los desinfectantes a base de amonio cuaternario (Roccal-D), fenol, hidróxido sódico o agentes oxidantes son eficaces en la inactivación del PCV-2 in vitro. Dada la importancia económica del PCV-2, las estrategias efectivas de manejo, detección temprana de infecciones desempeñan un papel crucial en la prevención de la propagación del PCV-2 y el control son esenciales para mitigar el impacto de la enfermedad en la industria porcina.
- Distribución geográfica de la infección por PCV-2
Siendo un virus ubicuo presente en prácticamente todas las granjas de cerdos. Es importante destacar que estos carecen de una distribución geográfica global similar y no todos inducen signos clínicos graves en los cerdos. PCV-2a, PCV-2b y PCV-2d se consideran los genotipos primarios debido a su alta prevalencia.
Se ha sugerido como un factor contribuyente a través de la selección de variantes de escape inmunológico. Investigaciones anteriores de filoevolución de PCV-2 han demostrado que los principales genotipos han surgido en oleadas periódicas, donde se establecen en camadas, juego aumentan su prevalencia entre las poblaciones porcinas. Esta distribución mundial de PCV-2 se debe a la intensificación de la producción porcina y al comercio internacional de porcinos, los cuales en caso de estar infectados contribuyen a diseminar el patógeno. El movimiento de cerdas de determinadas líneas genéticas para centros de producción de porcino, los cuales son la base del tejido productivo, se considera el principal motivo de la rápida expansión del PCV-2 y de los cambios en sus genotipos predominantes cada cierto tiempo.
- Circovirosis Porcina (PCV)
El circovirus que infectan a los porcinos (PCV) taxonómicamente se incluyen en el género Circovirus de la familia Circoviridae, “Circo” indica que el genoma viral tiene una conformación circular y el género Gyrovirus (“Gyro” deriva del griego gyrus que significa anillo o circuito). El genoma codifica un número limitado de proteínas, incluida la proteína replicasa (Rep) y la proteína de la cápside (Cap). También existen varias proteínas accesorias codificadas por el genoma viral.
El PCV-2 es virus no envuelto, con una cápside de simetría icosaédrica, de aproximadamente 1.76 kilobase (kb; es una unidad de medida que se usa como ayuda para designar la longitud del ADN). La proteína de la cápside forma la capa exterior del virión y encapsula el genoma viral, el que protege a una cadena simple o de una sola cadena de ADN circular (ssDNA, por sus siglas en inglés: single-stranded DNA), con un diámetro de alrededor de 17 a 20 nanómetros, la cual está compuesta por entre 1766-1768 nucleótidos de longitud de polaridad negativa: Este número tan reducido de nucleótidos y la ausencia de envoltura externa, hace que sea considerado de los virus de replicación autónoma más pequeños que se conocen.
Los circovirus se describieron originalmente como especies aviares, pero algunos de ellos se han identificado en peces, insectos y mamíferos. Hasta el momento se han descrito cuatro especies de circovirus que afectan a los cerdos:
- Circovirus tipo 1 (PCV-1). PCV-1 se describió por primera vez en 1974 y fue considerado no patogénico en cerdos.
- Circovirus tipo 2 (PCV-2).
- Circovirus porcino tipo 3 (PCV-3).
- Circovirus tipo 4 (PCV-4).
El PCV-2 es un virus que presenta una elevada especificidad de hospedador, siendo los cerdos el hospedador natural, haciendo que PCV-2 sea uno de los patógenos más importantes del porcino, aunque también se ha detectado en jabalíes, y en los pecaríes, aunque sólo los dos primeros pueden desarrollar alguna enfermedad asociada a la infección.
El PCV-3 puede contribuir a mayores pérdidas económicas que el PCV-2 por la fácil transmisión y propagación entre especies y a una alta tasa evolutiva. El control de los nuevos PCV es fundamental para disminuir la posibilidad de la propagación de varios genotipos en el futuro. Debido al aislamiento de PCV-3 de cerdos con problemas reproductivos, es probable que haya un vínculo entre el PCV-3 y el fracaso reproductivo. Los estudios retrospectivos de PCV-2 mostraron que PCV-2 estaba presente en la población de cerdos antes de que aparecieran problemas clínicos graves. Al igual que con el PCV-2, la susceptibilidad de los cerdos al PCV-3, el patomecanismo de la infección por PCV-3 y otros factores pueden cambiar y hacer que la infección por PCV-2 sea más grave.
| Tabla 4.1. Principales características de los circovirus porcinos | ||||
| PCV-1 | PCV-2 | PCV-3 | PCV-4 | |
| Año de descubrimiento | 1974 | 1997 | 2016 | 2019 |
| Distribución | Mundial | Mundial | Mundial | China
España |
| Prevalencia | Baja | Alta | Alta | Desconocida |
| Nucleótidos en el genoma | 1758-1769 | 1766-1769 | 1999-2001 | 1770 |
| Diversidad genética | Baja | Alta
(8 genotipos) |
Media* | Desconocida |
| Patógeno en condiciones de campo | No | Si | Si | Si |
| Primer aislamiento en algún cultivo celular | 1974 | 1977 | 2020 | No exitoso |
* Se ha observado cierta diversidad genética, pero aún hay consenso para considerarlos como subtipos. Modificado de López-Lorenzo, (2021), y Arnal et al., (2024).
Finalmente, en el año 2019, se describió un nuevo circovirus, denominado PCV-4, que fue aislado en cerdos de final de la transición-inicio de engorda con signos clínicos respiratorios, digestivos y con el Síndrome de dermatitis y nefropatía porcina (PDNS, por sus siglas en inglés: porcine dermatitis and nephropathy syndrome), sin embargo, su patogenicidad está todavía por demostrarse. En la Tabla 4.1 se indican las principales características de cada uno de ellos.
- Organización del genoma
El tamaño del genoma de los circovirus porcinos varía dependiendo de la especie de circovirus, que va desde 1758 nucleótidos (nt) hasta 2001 nt. El genoma de PCV-2 está formado por una cadena simple de ADN circular, compuesta por entre 1766-1769 nt, con una organización ambisentido (contine segmentos tanto de sentido positivo como negativo) y en el que se han diferenciado hasta 11 regiones, o fragmentos de lectura (ORF, por sus siglas en inglés: Open reading frame ). Se denominan desde ORF1 hasta ORF11 (fig. 4.1), y de ellos destacan principalmente tres ORF, ubicados en hebras opuestas, ORF1 y ORF2, y ORF3.
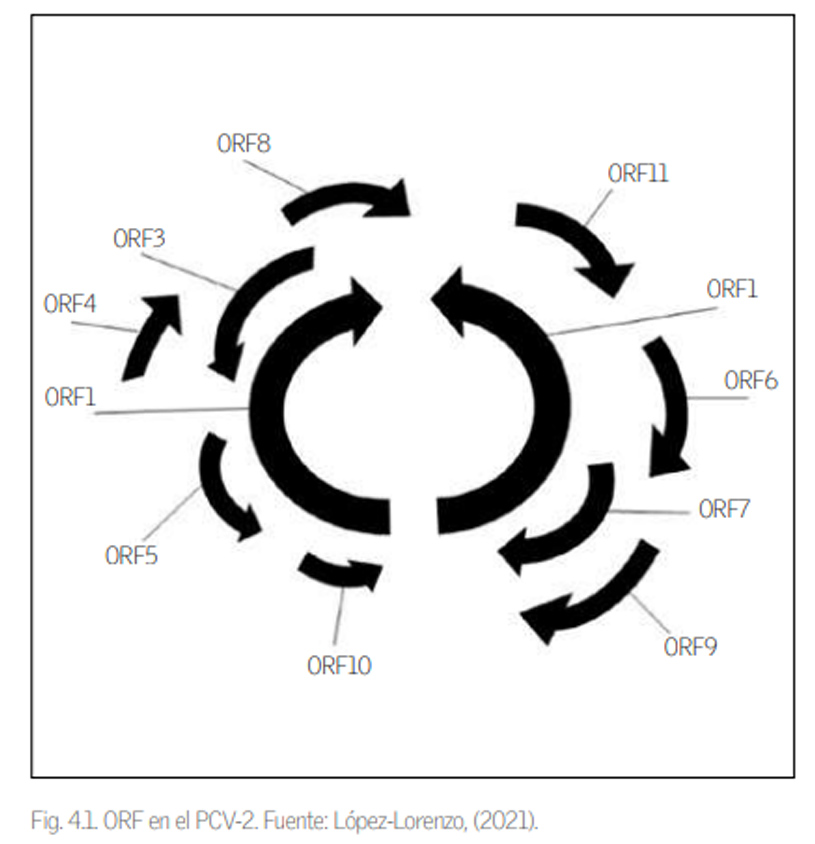
ORF1 es el más grande y el que menos variaciones presenta. Codifica la información necesaria para las proteínas encargadas de la replicación del virus, de las cuales destacan proteínas replicasa (Rep) 312 aminoácidos (aa) y Rep´ (178 aa). El ORF2 es el que codifica la información para la proteína de la cápside (Cap; 233 aa), la cual es la responsable de la formación de la cápside del virus. Esta cápside es el principal antígeno del PCV-2 y la que provoca la mayor respuesta inmune. Este ORF2 presenta variaciones, motivo por el que se utiliza en los estudios filogenéticos y es el responsable de la diferenciación entre genotipos de PCV-2.
El ORF3 está implicado en la apoptosis de las células infectadas por el PCV-2, que contribuye a la patogénesis viral, mientras que el ORF4 se opone al restringir la transcripción de ORF3 y evitando la apoptosis inducida por el PCV-2. Hasta la fecha, los demás ORF no están tan estudiados, pero se sabe que varios de ellos participan en la expresión de citoquinas e interleucinas en las células que infecta el virus, así como dando una función estructural.
- Patogenia
El PCV-2 tiene una alta prevalencia en las poblaciones porcinas de todo el mundo. Casi todas las piaras de cerdos están expuestas al virus y se puede encontrar en diversos tejidos y fluidos corporales de animales infectados y susceptibles. Los lechones se pueden infectar con el PCV-2 durante la vida prenatal o postnatal, dando lugar a diferentes manifestaciones clínicas de la enfermedad.
El virus se dirige principalmente a los tejidos linfoides, lo que provoca inmunosupresión y una mayor susceptibilidad a infecciones secundarias. Los mecanismos de patogénesis del PCV-2 implican la replicación viral, la modulación de la respuesta inmune del huésped y la inducción de la apoptosis.
- Embriones y fetos
Si la infección de las cerdas se produce durante la gestación es posible que el PCV-2 alcance a los embriones o a los fetos mediante transmisión transplacentaria. Los embriones son refractarios a la infección mientras estén recubiertos por la zona pelúcida. Una vez que ésta desaparece, el virus se puede replicar en la mórula y los blastocitos ocasionando la muerte y la reabsorción de los embriones.
Cuando la infección ocurre durante la fase fetal, la replicación del PCV-2 es mayor si el virus alcanza a los fetos de 40-70 días de gestación, el virus se replica principalmente en el corazón, seguido en menor medida en hígado, los órganos linfoides y los pulmones, a partir de ese momento son inmunocompetentes y la replicación es más reducida. En consecuencia, se produce un daño cardíaco que puede provocar la muerte y momificación del feto.
- Lechones y cerdos
Tras el nacimiento, el tropismo celular (la capacidad de un virus para infectar un tipo específico de célula o tejido) se centra en linfoblastos y células de la línea celular monocítica, los primeros son totalmente susceptibles mientras que en las células del linaje monocítico, la presencia de partículas virales rara vez conduce a una infección productiva. En los lechones y los cerdos adultos se considera que la replicación ocurre en tejidos linfoides más próximos a la vía de entrada del virus. No hay consenso sobre el tipo celular en el que el PCV-2 se replica. Algunos autores sostienen que las células diana son los linfocitos, mientras que otros sostienen que son células de la línea monocito-macrófago.
Además, el PCV-2 también se puede replicar en los linfocitos B y T sanguíneos. Esto, junto a la capacidad del virus de persistir en células dendríticas sin perder poder infectante, facilita su diseminación por todo el organismo y posterior replicación en otros tejidos linfoides, en células del epitelio bronquial y bronquiolar, del epitelio renal, epitelio del colon, células endoteliales, hepatocitos y células glandulares gástricas y pancreáticas.
De esta forma, el PCV-2 se puede detectar en prácticamente cualquier órgano de cerdo infectado, aunque, debido a su preferencia por células linfocitarias, los órganos linfoides son en los que se detecta la mayor cantidad. De hecho, se ha estimado que la concentración de PCV-2 en los ganglios linfáticos es entre 102 y 104 veces superior a la detectada en la sangre. Por tanto, el daño que provoca en prácticamente todos los sistemas del organismo hace que el cuadro clínico sea muy amplio.
- Transmisión
El PCV-2 tiene la habilidad de transmitirse a través de diferentes vías, sin embargo, la infectividad depende de factores como la susceptibilidad del huésped. El PCV-2 exhibe diversidad genética y se han identificado diferentes genotipos y subtipos. La diversidad genética del PCV-2 está asociada con variaciones en la patogenicidad, así como la forma y tipo de exposición, así como en la distribución geográfica. Ser un virus que se considera presente en prácticamente todas las granjas, que alcanza elevados porcentajes de infección en casos sin vacunar, y que tras años de vacunación no se ha logrado erradicar en ninguna granja comercial, confirman que el PCV-2 se transmite de forma muy eficiente.
En general, se ha estimado que la ratio de reproducción (R0) para esta infección es de 5.9 y que el tiempo medio de generación de enfermedad (dT; [tiempo para que un cerdo recién infectado contagie a otro susceptible]) es de 18.4 días. Sin embargo, estos parámetros oscilan ligeramente dependiendo del grado de contacto entre cerdos y de los días tras la infección. En condiciones experimentales, el R0 entre cerdos en una misma cuadra alcanza un valor de 8.9 mientras que entre animales en distintos corrales se reduce a 1.2; además, la transmisión es más eficiente entre los 8 y los 15 post-infección, disminuyendo a partir de este momento, aunque aún pueden permanecer infectantes hasta los 42 días post-infección.
Los estudios de infección experimental han demostrado que el PCV-2 es excretado a través de diferentes secreciones tanto de animales enfermos como de cerdos con infección subclínica, pero la cantidad de virus eliminado es mayor en animales con signos clínicos asociados al PCV-2. Este virus se excreta en gran cantidad y por numerosas vías, habiéndose detectado mediante la reacción en cadena de la polimerasa (PCR) en los fluidos indicados en la fig. 6.1. Esta excreción por múltiples vías en los parámetros mencionados en el párrafo anterior, pues la contaminación de superficies en las que están en contacto cerdos alojados conjuntamente es una vía que contribuye a su contagio.

En cerdos que no han sido vacunados frente a PCV-2, el pico de excreción se alcanza aproximadamente a los 15 días post-infección y desciende significativamente a partir de los 35 días. No obstante, esta excreción puede ser muy duradera, habiéndose detectado en muestras de leche hasta el día 27 de lactancia, en semen hasta 47 días post-infección y en heces e hisopos nasales desde los 28 hasta los 209 días de vida. También se ha visto que la excreción es mayor en los cerdos con PCV-2-SD o CP, que en los infectados de forma subclínica; en cambio, en cerdos vacunados la excreción es más reducida y de menor duración.
- Transmisión horizontal
El PCV-2 pueda excretarse por varias rutas, además de tener gran resistencia en el ambiente, hace que su transmisión horizontal sea eficiente. Se considera que las vías naso-oral y la fecal-oral son las principales vías de transmisión del PCV-2. La principal forma de transmisión de PCV-2 es el contacto directo entre cerdos infectados y susceptibles. Se ha visto que el contacto entre ambos favorece la transmisión de la infección y la aparición de signos clínicos, siendo más eficiente si los cerdos están en contacto directo que si se interpone algún elemento físico.
Se ha sugerido que la excreción de PCV-2 a través de la leche podría ser el origen de la infección que ocurre tras el descenso de la inmunidad maternal. Esta hipótesis se basa en que el PCV-2 quedaría latente en el organismo del lechón y se reactivaría una vez que la inmunidad maternal desciende. Se ha comprobado que, al menos en fetos, las células del timo pueden estar infectadas de forma latente. No obstante, esta hipótesis no se ha confirmado, pues se comprobó que tras un destete precoz (a los 10-12 días) y el traslado de esos lechones a condiciones controladas, estos no se infectaron hasta las 15 semanas, cuando finalizó el estudio.
La excreción de PCV-2 a través del semen es intermitente y, en base a la información hasta la fecha, insuficiente para infectar a las cerdas o a los embriones mediante inseminación artificial; no obstante, podría tener repercusión en casos de monta natural, pues la concentración viral es mayor que en las dosis seminales. El hecho que prácticamente todos los animales se infectan en un momento u otro a lo largo de su vida indica que la transmisión horizontal del PCV-2 es por lo tanto muy eficiente.
- Transmisión indirecta
Se ha detectado PCV-2 en el aire de las granjas y sugieren que la transmisión aerógena del PCV-2 puede ser factible al inhalar una cantidad elevada de PCV-2. Este hecho se confirma debido a que cerdos sin signos clínicos enfermaban al estar expuestos al aire procedente de salas con cerdos con PCV-2-SD.
Es probable que la transmisión entre especies de los PCV sea una grave amenaza para la industria porcina y otras industrias animales. La contaminación de los piensos con productos de pollo y porcinos infectados por PCV es una fuente potencial de infección para los animales de granja. Se detecto PCV-2 en muestras de médula ósea, sangre y cerebro de humanos, ratones, vacas y terneros. Este virus también se determinó en moscas domésticas y mosquitos en granjas de cerdos. Recientemente, se documentó la primera aparición de PCV-2 en mapaches y zorros.
Los roedores merecen una mención aparte porque, en condiciones experimentales, se han infectado e incluso se ha llegado a transmitir la infección entre ratones. No obstante, no hay consenso en cuanto a esta información, ya que otros estudios mostraron resultados opuestos. En condiciones de campo se ha detectado ADN de PCV-2 en muestras de roedores presentes en granjas de cerdos, no así en los capturados en los alrededores, aunque nunca se ha demostrado que puedan transmitir PCV-2 a los cerdos. También se ha detectado ADN de PCV-2 en moscas de granjas de cerdos, las cuales se sabe que no son susceptibles de infectarse, pero sí pueden portar el virus.
- Transmisión vertical
La transmisión vertical de cerdas a lechones a través de la inoculación in útero de fetos por medio de una infección transplacentaria, o mediante la infección intranasal de cerdas preñadas antes del parto. La progresión de los signos clínicos de la enfermedad en las cerdas depende de una alta replicación viral o de una viremia prolongada y del momento de la infección viral, que también puede determinar la aparición de fallo reproductivo clínico, también contribuye a la propagación del virus. En condiciones de campo se ha notificado el nacimiento de lechones virémicos procedentes de cerdas también infectadas; además, parte de esos lechones eran seropositivos y otros seronegativos, indicando que la infección se puede producir antes y después de que los fetos sean inmunocompetentes. En los casos en que se produce la transmisión por vía vertical, lo habitual es observar alteración reproductiva a nivel de granja, como un incremento de lechones nacidos muertos, momificados o camadas reducidas. En camadas sin alteraciones, se ha visto que el nacimiento de lechones infectados es prácticamente testimonial.
- Replicación vírica
Se ha visto que el PCV-2 infecta diversos tipos celulares, el virus o antígeno del virus ha sido aislado o detectado en órganos linfoides, hígado, riñón, placas de Peyer del intestino, pulmón, tonsila y otros tejidos, incluido el sistema nervioso central. El PCV-2 utiliza como receptores celulares los glicosaminoglucanos de heparán sulfato y condroitín sulfato presentes en la membrana de las células. A continuación, se transfiere hacia el citoplasma celular a través de un mecanismo de endocitosis. Una vez en el citoplasma, el PCV-2 se libera de la endosoma mediante mecanismos que van desde una acidificación hasta un bloqueo en el cambio de pH, dependiendo del tipo de célula infectada. Posteriormente, el ADN del PCV-2 se transporta al núcleo de las células, donde las Rep, Rep´y Cap son transcritas y las proteínas son sintetizadas e importadas desde el citoplasma. Posteriormente tiene lugar la elongación del ADN del PCV-2 y el ensamblaje del virus, acciones para las que sirve de enzimas de la célula hospedadora presentes en la fase S del ciclo celular. Finalmente, se liberan los viriones e infectan a otras células susceptibles siguiendo el mismo mecanismo.
El ácido nucleico o antígenos de PCV-2 se encuentra generalmente en el citoplasma de histiocitos, células gigantes multinucleadas y otras células de la línea monocito/macrofágica tales como macrófagos alveolares, células de Kupffer y células dendríticas foliculares de los tejidos linfoides. También es posible encontrar ácido nucleico o antígeno viral en el citoplasma del epitelio renal y respiratorio (bronquial, bronquiolar y alveolar), endotelio vascular, células acinares y ductales pancreáticas, y en el núcleo de células de la línea monocito/macrofágica, células musculares lisas, hepatocitos, y enterocitos. Además, se ha detectado PCV-2 en neuronas, en células mononucleares del infiltrado perivascular cerebral, células similares a fibroblastos, células endoteliales en meninges, plexos coroideos, cerebro y cerebelo de cerdos infectados, principalmente en el citoplasma y ocasionalmente en el núcleo. En los fetos, la principal célula que contiene PCV-2 es el cardiomiocito, tanto en el núcleo como en el citoplasma. Dentro de las células de los tejidos de cerdos infectados pueden verse cuerpos de inclusión intracitoplasmáticos intensamente basófilos de 5 a 25 nm de tamaño.
Sin embargo, no se sabe cómo el PCV-2 establece la infección y se replica en los cerdos, pudiéndose replicar “in vitro”, en células PK-15, en la línea celular Vero de riñón de mono verde africanos, en células semicontinuas de pulmón porcino, en células semicontinuas de testículo porcino, etc. En los cultivos celulares, el virus no causa lisis visible de las células infectadas, y su presencia puede demostrarse solo por métodos indirectos, tales como inmunofluorescencia, inmunohistoquímica, hibridación in situ. Usando estas técnicas, el PCV-2 sólo puede encontrase en un pequeño número de células, pudiendo presentar cuerpos de inclusión citoplasmáticos heterogéneos, consistentes en redes electrodensas paracristalinas de pequeñas partículas víricas sin envoltura.
- Leucopenia y objetivos celulares asociados a la infección por PCV-2
La alteración del sistema inmune es la principal característica en los cerdos infectados por PCV-2. A causa del daño provocado en los órganos linfoides, es posible que en estos se reduzca el número de linfocitos, provocando una depleción linfoide. Esta depleción linfoide depende de la cantidad de PCV-2 en el tejido linfoide: a mayor cantidad de virus, mayor depleción. A consecuencia de esta alteración surge una linfopenia (1. Linfocitopenia, 2. Reducción en la cantidad de linfa, Linfocitopenia: reducción en el número absoluto de linfocitos por unidad de volumen de sangre periférica), detectable a los 7-10 días post-infección. Esta linfopenia afecta principalmente a los linfocitos de la línea T, y en menor medida a los de la línea B, y está acompañada de una disminución de células Natural Killer (NK) motivos por los que en los cerdos con PCV-2-SD la respuesta inmune celular está disminuida. No obstante, aquéllos que sobreviven a la infección por PCV-2 desarrollan inmunidad frente al virus.
La leucopenia en el caso de cerdos que están desarrollando PCVD, es detectable antes de la aparición de los síntomas clínicos. Este tipo de cinéticas ofrecen cierto valor como marcador diagnóstico útil para animales que van a desarrollar la enfermedad. Las células con las cuales están más frecuentemente asociados el PCV-2 en animales infectados, y en las que el virus puede hallarse durante periodos prolongados después de la infección, son las monocíticas (monocitos, macrófagos y células dendríticas; [DC, por sus siglas en inglés: Dendritic Cells]). Esta presencia anormal y prolongada del virus en las DC es un importante problema para el sistema inmunitario, por el papel crítico de las células como piezas centrales en el desarrollo y regulación de la defensa inmunitaria.
Por un lado, las DC son importantes para la defensa inmunitaria innata, en su papel de vigilantes de las superficies mucosas del hospedador. Allí, dichas células pueden responder a los patógenos, alergenos y otras “señales de peligro”, que podrían suponer una amenaza para el hospedador. Además, las DC controlan el desarrollo de respuestas antígeno específicas, que ofrecen al hospedador protección prolongada contra un patógeno en concreto, como ocurre después de una vacunación eficaz. Por esta última razón, se alude a ellas como células presentadoras del antígeno “profesionales”. Dichas estructuras también están implicadas en el control de la supervivencia y el funcionamiento de los linfocitos en el cuerpo (homeostasis).
Cuando los patógenos modifican la función de las DC, se puede dar lugar a que el reconocimiento del “peligro” se vea alterado. Esto debilitará la defensa inicial contra el ataque de patógenos, así como la capacidad de las DC para controlar actividades inmunológicas. La interferencia con éstas últimas conducirá a respuestas inflamatorias excesivas, sobreactivación de linfocitos o alteración de la homeostasis.
Por las razones descritas, la presencia de PCV-2 en las DC supone una amenaza para la capacidad del hospedador de responder frente a los patógenos y resistir ante muchos de ellos, así como ante otros “peligros” a los que se enfrenta regularmente. Además, los animales infectados con PCV-2 presentan antígenos virales en macrófagos y DC, células que, pese a la infección, permanecen viables, implica una persistencia del virus en estas células y en el hospedador. Los análisis in vitro han confirmado la predilección del PCV-2 por las DC y los macrófagos. Curiosamente, estas células acumulan antígenos virales durante periodos prolongados, sin que haya evidencia clara de una replicación del virus. De acuerdo con esto, las DC y los macrófagos pueden considerarse portadores de PCV-2 (virus vivo, antígenos y ADN virales), y por ello desempeñan un papel fundamental en la persistencia y diseminación de este patógeno en las piaras y desde las madres a los lechones.
Por supuesto, tiene que haber una fuente de virus en replicación para que las DC y los macrófagos acumulen niveles tan elevados de PCV-2. Los estudios in vitro han demostrado que este patógeno infectará y se replicará tanto en las células endoteliales como epiteliales del intestino. La observación de que la replicación viral se pone en marcha cuando las células se activan y comienzan a dividirse. Este tipo de acciones pueden ocurrir durante el desarrollo de respuestas inflamatorias locales y en lugares de daño tisular.
- La infección de las células dendríticas por PCV-2
El PCV-2 no induce la muerte en las DC que infecta, ni tampoco en los linfocitos asociados. De acuerdo con esto, la linfopenia observada in vivo probablemente sea un efecto indirecto de la infección por PCV-2 en las células mielomonocíticas. Por inferencia respecto a otros virus monocitotrópicos, esta asociación de PCV-2 con células monocíticas podría explicar la leucopenia, debido a una emisión de señales aberrantes que parte de las células infectadas y que interfiere con la homeostasis. Sin embargo, la presencia del virus en las DC no perjudica su interacción inmunobiológica con los linfocitos; no obstante, el mantenimiento de cantidades relativamente altas de antígeno de PCV-2 y ADN viral en dichas estructuras, y la retención de PCV-2 infeccioso durante varias semanas no puede dejar de tener consecuencias.
A pesar de que las DC pueden procesar algunos de los antígenos virales que portan, lo que conduce a la inducción de las defensas inmunitarias, las grandes cantidades de antígeno presentes en ellas tienen consecuencias para la función defensiva. Esto lo atestigua el retraso de varios días o semanas que se da antes de que se visualice una respuesta inmunitaria anti-PCV-2, en contraste con la respuesta frente a una infección concurrente como por ejemplo con parvovirus porcino (PPV, por sus siglas en inglés; Porcine Parvovirus). Esta última infección se establece mucho más rápidamente que la de PCV-2 en el contexto de una infección concomitante. Por tanto, la linfopenia (especialmente de linfocitos B) podría explicar la tardía respuesta anti-PCV-2, pero no afecta a la rápida respuesta anti-PPV que se observa. Además, los animales asintomáticos que no muestran signos de linfopenia también manifiestan un reconocimiento inmunitario de PCV-2 lento.
- Mecanismos de afectación de la competencia
Considerando las características previamente descritas de la infección por PCV-2 en las DC, la señal inmunomoduladora que proporciona el virus es tanto sutil como restringida. Por lo tanto, no fue sorprendente descubrir que el objetivo para la modulación de las defensas inmunitarias inducida por el PCV-2 se centra en la capacidad de las DC para reconocer “señales de peligro”. La rápida detección de un daño potencial por las defensas inmunitarias innatas es el objetivo de la inmunomodulación que provoca el PCV-2. Como se ha comentado anteriormente, estos procesos son críticos tanto para las defensas innatas como para las promociones de respuestas inmunitarias específicas. Así, resultan especialmente sensibles los procesos de reconocimiento de patógenos mediados por receptores tipo peaje (TLR, por sus siglas en inglés: toll-like receptos).
Las estructuras más sensibles dentro de las defensas inmunitarias innatas son las DC plasmacitoides, responsables de la producción del interferón y los factores de maduración del factor de necrosis tumoral, que son esenciales para la maduración de las DC mieloides. Aunque estas últimas todavía son capaces de responder a los factores de maduración, el perjuicio provocado por el PCV-2 sobre el factor de maduración de las DC plasmacitoides evitará de forma eficaz que se desarrollen respuestas inmunitarias eficientes frente a otros patógenos.
La situación se ve agravada por la observación de que la mayoría de las células presentadoras de antígeno y auxiliares pueden verse perjudicadas por la presencia de una infección por PCV-2 (hasta la fecha esta ha sido observado para el reconocimiento de los ligandos TLR2, TLR3, TLR7, TLR8 y TLR9). Esta interferencia con el reconocimiento de los “peligros” tiene serias consecuencias en términos de alterar el reconocimiento rápido y eficiente de diversos virus y bacterias, lo cual favorece el establecimiento de sus infecciones concomitantes junto con los consiguientes síntomas y enfermedades. Algunas pruebas mostraron que este efecto modulatorio del PCV-2 está agravado por las DC mieloides infectadas, que producen factores que promueven una regulación a la baja, más que la activación de respuestas inmunitarias específicas.
Curiosamente, el componente viral más efectivo para provocar tales señales moduladoras es la forma replicativa circular de doble cadena del genoma del ADN viral. Resulta importante que, al parecer, este ADN bicatenario debe alcanzar unos niveles mínimos dentro de las DC antes de que se vea comprometida la funcionalidad de éstas. Esto puede explicar por qué sólo ciertos animales desarrollan los síntomas de la enfermedad. Además del ADN de cadena doble, previas pruebas probaron que la cápside proteica del virus puede interactuar con varios componentes celulares, pero aún no se ha determinado si esto modulará el desarrollo de las defensas inmunitarias, como ocurre con la forma replicativa del ADN viral de doble cadena.
![Granja Porcina: [Guía básica] para iniciar con la tuya](https://blog.deacero.com/hs-fs/hubfs/C%C3%B3mo%20comenzar%20mi%20granja%20porc%C3%ADcola%20-%20panel%20porcicola.jpg?width=4217&name=C%C3%B3mo%20comenzar%20mi%20granja%20porc%C3%ADcola%20-%20panel%20porcicola.jpg)
Deja un comentario